





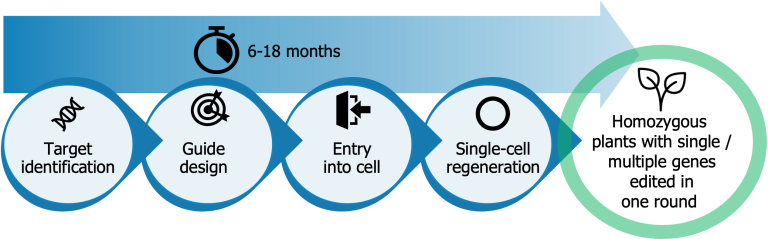
The key strength of CRISPR-based breeding is that it allows for faster and more targeted development of crop varieties. After several years of development, CRISPR has become a mature enough technology to deliver the plants that we need for the future, in a safe and controlled manner. Where similar type of crop adjustments would traditionally take between seven and ten years (if possible at all), they can now be done within two to four years.
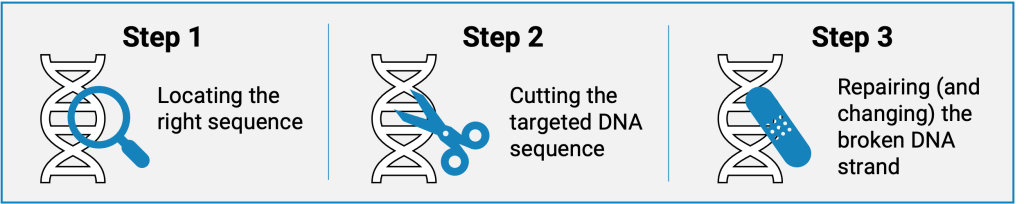
To change a specific DNA sequence, the CRISPR system relies on a DNA cutting enzyme (endonuclease) that is guided to a target sequence using a guide RNA (gRNA). By cutting and later repairing the DNA, the sequence can be changed in a desired way. 3 main steps are necessary:
It has been a journey developing the plant glycan knock-out cell line. The team at HRB, who are real experts of the TiGER technology, could translate it to our needs and cared about us as clients. Their service helps us to achieve our goal to bring new pharmaceuticals to patients that are effective and affordable, wherever they are needed.
